
FAQ

WHAT DOES THE NOMENCLATURE OF STRAINS MEAN?
OUTBRED STOCK
1) Laboratory Code holding the stock
2) Laboratory Code holding the original stock
3) The two points “:”
4) Strain symbol
Example: SPRAGUE DAWLEY RjHan: SD = Rj for Janvier Labs + Han for Hannover + the two points + SD for Sprague Dawley.
INBRED STOCK
1) Strain symbol
2) Slash “/”
3) Laboratory Code holding the original stock
4) Laboratory Code holding the stock
Example: BROWN NORWAY BN/OrlRj = BN for Brown Norway + slash + Orl for Orléans CSAL + Rj for Janvier Labs.
MUTANT STOCK
- Oubred mutant stock:
1) Laboratory Code holding the stock
2) Strain symbol
3) Gene + mutation
Example: RjNMRI–Foxn1nu/nu = Rj for JANVIER LABS + NMRI for the strain + Foxn1 for the gene + nu/nu for the mutation on the gene.
- Inbred mutant stock:
1) Strain symbol
2) Laboratory Code holding the original stock
3) Laboratory Code holding the stock
4) Gene + mutation
Example: Balb/cAnNRj–Foxn1nu/nu = Balb/cAn for the strain + N for NIH + Rj for JANVIER LABS + Foxn1 for the gene + nu/nu for the mutation on the gene.
Except for the obese and diabetic mice nomenclature:
1) Strain symbol
2) Gene + mutation
3) Slash “/”
4) Laboratory Code holding the original stock
5) Laboratory Code holding the stock
Obese B6.V-Lepob/JRj;
Diabetic BKS(D)-Leprdb/JOrlRj
HYBRID STOCK
1) Abbreviated strain symbol for females
2) Abbreviated strain symbol for male
3) F1
4) Slash “/”
5) Laboratory Code holding the original stock
6) Laboratory Code holding the stock
Example: B6CBAF1/JRj = B6 for female C57BL/6J + CBA for male CBA/J + F1 for the first generation of the cross + slash + J for The Jackson Laboratory + Rj for JANVIER LABS.
Other abbreviated strain symbol: D2 for DBA/2J, SJL for SJL/J, C for BALB/cJ, C3 for C3H
LABORATORY REGISTRATION CODE OR LABORATORY CODE
Laboratory Registration Code or Laboratory code identifies a particular institute, laboratory, or investigator that produced, and may hold stocks of, for example, a DNA marker, a mouse or rat strain, or were the creator of a new mutation. Laboratory codes can be assigned through MGD (or MGI), RGD or directly by the ILAR.
- Rj: JANVIER LABS (Centre d’élevage Roger Janvier)
- J: The Jackson Laboratory
- N: National Institute of Health (NIH)
- Han: Hannover – Zentralinstitut fur Versuchstierzucht
- He: W. E. Heston
- Orl: Orléans CSAL – Institut de transgenose TAAM UPS44
- Ibmm: Institut de Biologie et de Médecine Moléculaire (Universite libre de Bruxelles)
- Icr: Institute for Cancer Research
- Tub: Tumblebrook Farm (Tum)
- Alhn: Agnes Lehuen (INSERM U25, hospital Necker)
- Lpf: Françoise Lepault (INSERM U561, hôpital St Vincent de Paul)
- Pas: Institut Pasteur
- Kyo: Kyoto University
Accede to Institute for Laboratory Animal Research #1 ILAR or #2 ILAR
Accede to Mouse Genome Informatics (MGI)
Accede to Rat Genome Database (RGD)
WHAT ARE THE DIFFERENCES BETWEEN STRAINS?
For a study, the first point is to know whether outbred mice or inbred mice will be used:
– Outbred strains are genetically undefined; that is, no two individuals from an outbred stock are the same. The purpose of an outbred stock is to maintain maximum allele variability. Outbred strains have genetic heterogeneity and is characterized by allele frequency in their population. This characteristic makes outbred colony able to mimetic variable response in the same way as observed in random groups like observed in human population. The individual genotype is not reproducible but the allele distribution in the colony is typical of the colony. Phenotypic variation depends on interaction between genes and the environment.
– Inbred strains are homozygotes on almost all loci. They share the same genome, this removes inter-individual genetic variability.


The C57BL/6JRj and the C57BL/6NRj differ in their origin, genetics, behaviour and physiology:
ORIGIN
The C57BL strain, inbred strain, was isolated and identified by CC Little from a A. Lathrop’s stock crossing (1921). He gave up one of his colonies to the Jackson laboratory in the early 1930s. The colony of C57BL from The Jackson Laboratory was divided into sub-lines before 1937 giving rise to the C57BL/6 and the C57BL/10.
- The C57BL/6J strain was introduced at Jackson Laboratory in 1948 (F22). Janvier Labs acquired the C57BL/6J strain, from CSAL (Orleans) in 1993 at F172;
- The C57BL/6 strain was transmitted at the 32th generation (F32) from the Jackson Laboratory to the National Institute of Health (N.I.H) in 1951. A subsequent major discovery revealed that the transmitted strain had mutated giving rise to the B6N strain (C57BL/6N). Janvier Labs acquired the C57BL/6N strain from the National Institutes of Health (USA) in 1999.
GENETIC
- 34 SNPs, 2 indels and 15 structural variants were identified which distinguish the C57BL/6J from the C57BL/6N (Simon et al. 2013).
- Studies show differences in the genetic polymorphism between the C57BL/6J and the C57BL/6N (Bothe et al. 2004; Tsang et al. 2005; Mekada et al. 2009; Zurita et al. 2010).
- The C57BL/6N has not the deletion in the Nnt gene (nicotinamide nucleotide transhydrogenase), which is find in the C57BL/6J (Freeman et al., 2006).
- The C57BL/6N has the rd8mutation (retinal degeneration 8), not find in the C57BL/6J (Chang et al. 2002; Aleman et al. 2011).
BEHAVIOUR
Differences exist in the behaviour between the C57BL/6J and the C57BL/6N (Matsuo et al. 2010):
- Preference for ethanol, conditional fear, motor coordination and pain sensitivity (Blum et al. 1982; Bryant et al. 2008);
- Locomotor activity is higher and anxiety is lower in the C57BL/6J;
- Inhibition of terror is high in the C57BL/6N. The fear responses differ between C57BL/6J and C57BL/6N (Radulovic et al. 1998; Stiedl et al. 1999).
PHYSIOLOGY
Anesthetic effects on heart function differ between C57BL/6J and C57BL/6N (Roth et al. 2002) and the electroconvulsive threshold differs (Yang et al. 2003).
In the choice of the BALB/c strain, here are the differences between the 3 in our catalogue:
| BALB/cJRj
|
BALB/cAnNRj
|
BALB/cByJRj
|
|
| Origin | In 1935, Dr Snell sent BALB/c breeders to Dr Scott who brought them to The Jackson Laboratory in 1945. The strain became BALB/cJ in 1947 by The Jackson Laboratory acquisition. | In 1935, breeding stock was also provided by Dr Snell to Drs Heston and Andervont at the National Institute of Health (NIH). The Andervont subline (BALB/cAn) was incorporated into NIH main colonies in 1951 (BALB/cAnN). | In 1961, Dr Bailey acquired a colony of BALB/c mice at NIH. The BALB/cBy strain was created by him from the BALB/cAnN strain. When he moved to The Jackson Laboratory in 1967, he brought the strain (BALB/cBy) with him. In 1974 some of the breeding stock was transferred to the Production Department of The Jackson Laboratory at F136 and the J designation was added (BALB/cByJ). |
| Haplotype | H2d | H2d | H2d |
| Gene | Qa2a | Qa2b | Qa2b
Acadsdel-J (acyl-Coenzyme A dehydrogenase, short chain) Ahrb-2 (aryl-hydrocarbon receptor) Cdh23ahl [cadherin 23 (otocadherin)] |
| Phenotype | Aggressive Very active |
Superior yield in ascites production Good breeder with better performance than BALB/cJRj and BALB/cByJRj mice Less aggressive Active |
Superior yield in ascites production Less aggressive Very active |



Although these 3 strains are considered matched at the Major Histocompatibility Complex (MHC) (they have the H2d haplotype) and tissues can be transplanted between strains, the class 1b locus in BALB/cAnN and BALB/cByJ contains a deletion in Qa2 on Chromosome 17 (Mellor et al. 1985). The genes controlling the Qa2 lymphocyte alloantigen was discovered by Lorraine Flaherty and her colleagues (Flaherty et al. 1985). These genes are a cluster of class I-like genes located to the right of H2d in the mouse major histocompatibility (MHC) complex.
The different BALB/c strains can react differently at the physiological, behavioral and pharmacological levels.

The C3H parent strain was developed by LC Strong in 1920 from a cross of a Bagg albino female with a DBA male. In 1930, Strong sent a colony to Andervont who in turn sent one to Heston (at generation F35), who gave the name of C3H/He. Heston stock was then transferred to the NIH in 1951 (generation F51) which gave the name of C3H/HeN.
- Janvier Labs acquired the C3H/HeN from the NIH in 1996 (F169) which gave the name of C3H/HeNRj.
- The C3H/He strain was transmitted to the Jackson Laboratory in 1952 which gave the name of C3H/HeJ.
Thus, the C3H/HeJ and C3H/HeN have the same origin.
But these 2 strains differ at these following points:
- The C3H/HeN strain is Tlr4lps-n (toll-like receptor 4; lipopolysaccharide normal response) or formerly LPSn. This mouse strain will therefore respond normally to LPS stimulation that is why it is called LPS sensitive. In contrast, the C3H/HeJ is Tlr4LPS-d or LPSd and it doesn’t respond to LPS. The response to endotoxin LPS is subjected to a genetic control. This control is bound to the protein TLR4 (Toll -like receptor 4) formerly known as LPS.
- The alopecia observed in C3H/HeJ strain (in Janvier Labs, we don’t observe the alopecia phenomenon in our C3H/HeNRj mice):
- Sundberg et al, 1994, Alopecia Areata in Aging C3H/HeJ Mice, J Invest Dermatol 102:847-856:
Alopecia developed diffusely or in circular areas on the dorsal surface. Pedigree tracing of affected C3H/HeJ mice suggests that this non-scarring alopecia may be an inherited disease. This is a complex polygenic disease with a female predominance at younger ages. Female mice developed the disease earlier than male mice (3-5 vs > 6 months), with equal numbers affected by 18 months of age.
- Sundberg et al, 1994, Alopecia Areata in Aging C3H/HeJ Mice, J Invest Dermatol 102:847-856:
-
- Sun et al, 2008, The C3H/HeJ mouse and DEBR rat models for alopecia areata: review of preclinical drug screening approaches and results, Exp Dermatol. 2008 Oct; 17(10): 793–805:
The C3H/HeJ inbred mouse strain spontaneously develop adult onset alopecia areata (AA), a cell mediated disease directed against actively growing hair follicles.
- Sun et al, 2008, The C3H/HeJ mouse and DEBR rat models for alopecia areata: review of preclinical drug screening approaches and results, Exp Dermatol. 2008 Oct; 17(10): 793–805:
-
- Wikramanayake et al. 2012, Effects of the Lexington LaserComb on hair regrowth in the C3H/HeJ mouse model of alopecia areata, Lasers Med Sci. 2012 Mar; 27(2):431-6: Up to 20% of aged C3H/HeJ mice develop AA.
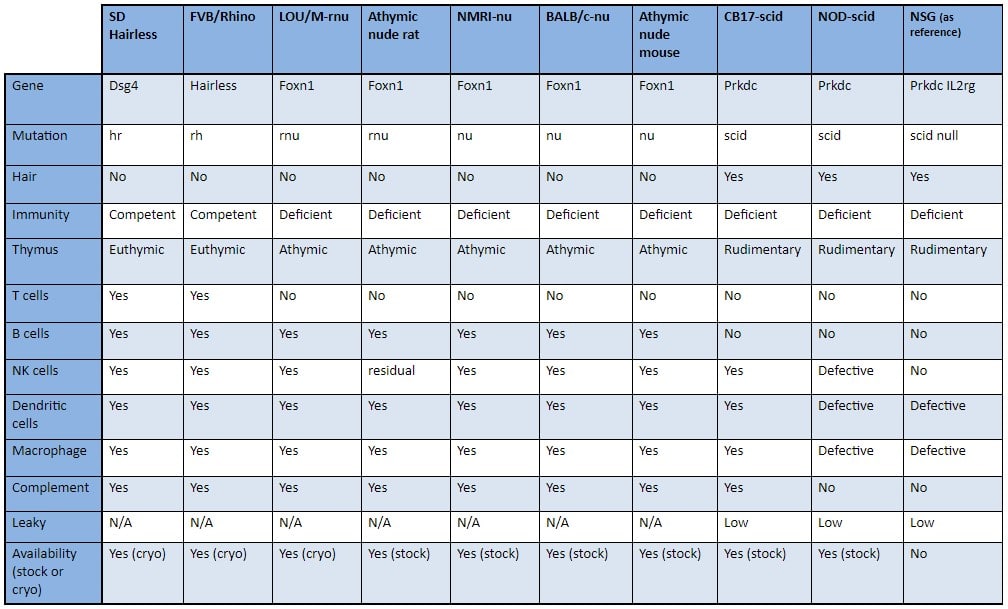
WHAT ARE THE CHARACTERISTICS OF SOME STRAINS?


Excellent breeder:
- Very productive
- Large litters (> 10)
- Very good parental qualities
- Example of strains: Sprague Dawley, Wistar, SWISS


Good breeder:
- Productive
- Medium litters (6-10)
- Good parent
- Example of strains: Dark Agouti, F344, SHR, FVB/N, NOD-scid

Challenging breeder:
- Not productive
- Little litters (< 6)
- Bad parental qualities (cannibalism, lactation defect, etc.)
- Example of strains: C57BL/6J, DBA/1, DBA/2, 129/Sv
Alopecia is a characteristic of the C57BL/6J strain.

The origin of this phenomenon remains uncertain:
- This was attributed to behavioral phenomena such overgrooming/barbering. Hair loss due to overgrooming (hair nibbling, whisker-eating) has been observed. In every case where overgrooming occurs the hair will grow back again if the mice are separated into individually occupied cages. If only one whisker-eating mouse is present in a cage, it can be readily identified by its own full set of whiskers
- Initially, this phenomenon has been associated with behavior “dominance” or of “aggression”
- Currently, the assumption changes and the track would move to an abnormal repetitive behavior (similar to OCD in humans)
- Genetic predisposition and environmental factors are involved
Alopecia begins shortly after weaning; it is growing around the age of 8-9 weeks and can reach a vast majority of animals of this age. The occurrence of hair loss, the size and shape of the affected area and the frequency of a particular pattern are variable. Mice of the C57BL substrains are much more prone to hair loss than other strains and certain congenic histocompatibility strains are especially susceptible. There may also be an individual variation.
Most often alopecia begins dorsally, with various degrees of damage. Some cases may progress to acute ulcer which can heal with a bed of granulation tissue, a pseudo-epitheliomatous hyperplasia and scarring. Others have varying degrees of itching that can lead to progressive ulcerative disease requiring euthanasia.
Thus, alopecia includes a slight focal alopecia, chronic ulcerative dermatitis and scarring alopecia which can be separated into:
- “Barbering”/trichotillomania for mild alopecia affecting areas of the head
- Focal alopecia present on the body that often begins on the dorsal skin behind the ears and may progress to chronic ulcerative dermatitis.
The body alopecia and ulcerative dermatitis are poorly understood. The data showed a predilection of this disease in females.
Hair loss caused by overgrooming is not a problem in our colonies. We are aware of the situation, the cause in not clearly understood, and the method of prevention (other than isolating the mice) is not known.
An alternative could be working on the C57BL/6N less sensitive to this phenomenon of alopecia (Cf. “Differences between C57BL/6J and C57BL/6N”).
References
- Sundberg et al. 2011, Primary follicular dystrophy with scarring dermatitis in C57BL/6 mouse substrains resembles central centrifugal cicatricial alopecia in humans, Vet Pathol. 48(2):513-24.
- Kalueff et al. 2006, Hair barbering in mice: implications for neurobehavioural research, Behav Processes. 71(1):8-15.
- Garner et al. 2004, Barbering (fur and whisker trimming) by laboratory mice as a model of human trichotillomania and obsessive-compulsive spectrum disorders, Comp Med. 54:216-24.
- Sarna et al. 2000, The Dalila effect: C57BL6 mice barber whiskers by plucking, Behav Brain Res. 108:39-45.
- the Jackson Laboratory : https://www.jax.org/news-and-insights/1987/october/alopecia-in-c57bl-6-and-related-mouse-strains
- Militzer and Wecker, 1986, Behavior associated alopecia areata in mice, Laboratory Animals 20:9-13.
- Strozik and Festing, 1981, Whisker trimming in mice, Laboratory Animals 15:309-312.
- Thornburg et al. 1973. The pathogenesis of the alopecia due to hair chewing in mice, Lab. Animal Sciences 23:843-850.
- Long, 1972, Hair-nibbling and whisker-trimming as indicators of social hierarchy in mice, Animal Behavior 20:10-12.

The teeth of rodents grow continuously. Eruption of the incisors in young mice and rats occurs between days 10-12 of age. In mice, the normal rate of eruption (which equals the rate of wear, so that the incisors remain a constant size in adult mice) is approximately 2mm/week for the upper incisors and 2.8mm/week for the lower incisors. This results in a turnover of the entire tooth in 35-45 days.
Rodent teeth wear naturally by eating hard foods and gnawing behaviour. In the laboratory, these conditions are mimicked by providing the animals with rodent diet formulated to be hard enough and materials to gnaw to wear the teeth.
Malocclusion should be the first disorder suspected when a mouse is smaller and thinner than its littermates at weaning. Malocclusion is a dental disorder of many strains of laboratory rodents and is readily diagnosed by an oral examination. It is a misalignment of teeth or incorrect relation between the teeth of the two dental arches. Dental malocclusion affects incisors of rodents, but may also affect the molars, resulting in impaired chewing. Due to a dental malformation, an animal has difficulty eating with discomfort to chew the food. This explains the low weight and bad state observed in these animals.
If the growing teeth penetrate facial structures, it may also result in oral and facial abscesses and osteomyelitis. The standard recommendation for animals with malocclusion is euthanasia. When the malocclusion is later in onset and due to trauma, oral tumor formation, or other causes, it is possible to manage the treatment of a valuable mouse through a program of regular tooth trimming using blunt-tipped scissors, to prevent inadvertent oral trauma, and adding a provision of powdered diet.
References
- Dontas et al. 2010, Malocclusion in aging Wistar rats, J Am Assoc Lab Anim Sci. 49: 22-6
- The Jackson Laboratory: https://www.jax.org/news-and-insights/2003/april/malocclusion-in-the-laboratory-mouse
- Petznek et al. 2002, Reduced body growth and excessive incisor length in insertional mutants mapping to mouse Chromosome 13. Mammalian Genome; 13:504-509.
- Peters and Balling, 1999, Teeth. Where and how to make them. Trends in Genetics; 15:59-65.
- Miller, 1977, Genetic traumatic occlusion in the mouse. J Periodontal Res; 12:64-72.
Mice vagina is closed at birth and open at the age of 24 to 28 days, around the onset of the sexual maturity.
The persistence of the vaginal septum is materialized by a septum or an imperforate vagina.
There may be serious health complications (distension of the vagina, cervix and uterus) for the mouse and inability to reproduce. If the mouse is mated despite this vaginal septum, its life may be endangered because it can cause difficulties during the birth (dystocia).
In C57BL/6J mice, the vaginal septum is well described:


This defect was identified in 1976, with incidence of 4.0% in this strain. A Study in 2004 (Gearhart et al.) reported incidence of 11.3%, nearly triple the original reported incidence.
For comparison, incidence of vaginal septum in C57BL/6N females was determined and was found to be 1%.
Imperforate vagina in the mouse is believed to be inherited in a complex recessive manner.
An alternative could be working on the C57BL/6N less sensitive to this phenomenon of vaginal septum (Cf. “Differences between C57BL/6J and C57BL/6N”).
References
- Gearhart et al. 2004, Increased Incidence of Vaginal Septum in C57BL/6J Mice Since 1976, Comparative Medicine, 54(4): 418-421
- Eisen, 1989
- Jubb et al.1985, Pathology of Domestic Animals. 3rd ed., Academic Press, Orlando
- Cunliffe-Beamer and Feldman, 1976, Vaginal septa in mice: incidence, inheritance, and effect on reproductive performance. Lab Anim Sci 26:895-898
- Green, 1975, Biology of the Laboratory Mouse, Dover Publications, New York.

Congenital hydrocephalus is usually observed in animals at weaning, or shortly thereafter, by the presence of a domed head, a consequence of deformation of the cranial box following hydrocephalus. The flow of cerebrospinal fluid is blocked leading to a build-up of fluid in the brain. This causes a swelling of the head and a compression of the brain tissue. However, hydrocephalus can occur at any time in life and the causes are multiple: intraventricular hemorrhage, head trauma, brain tumor, and venous hypertension.
Hydrocephalus may be discrete at first and phenotypic abnormalities such as bulging of the skull are not visible. Cases of hydrocephalus, discreet and morphologically unobtrusive, are found only during the examination of the brain with hypertrophy of the ventricles.
Hydrocephalus is described in the C57BL/6J strain (Hydrocephalus in Laboratory Mice, JAX® NOTES, July 2003: https://www.jax.org/news-and-insights/2003/july/hydrocephalus-in-laboratory-mice).
Hydrocephalus is observed and described in outbred rats as Wistar rats (Tu et al. 2014, Imaging of Spontaneous Ventriculomegaly and Vascular Malformations in Wistar rats: implications for Preclinical Research, J Neuropathol Exp Neurol. 2014 Dec; 73(12): 1152–1165).
WHAT ARE THE MAIN MUTATIONS FOUND IN MOUSE STRAINS?
Today, the CD45 (also named Ly5) gene is called Ptprc.
Protein tyrosine phosphatase receptor type c also known as PTPRC is an enzyme that is encoded by the ptprc gene (chromosome 1). Ptprc encodes CD45, a protein tyrosine phosphatase which is expressed in nucleated hematopoietic cells and is required for their activation. PTPRC is also known as CD45 antigen (CD stands for cluster of differentiation), which was originally called leukocyte common antigen.
CD45 is signalling molecules that regulate a variety of cellular processes including cell growth, differentiation, mitotic cycle, and oncogenic transformation. It has been shown to be an essential regulator of T- and B-cell antigen receptor (TCR and BCR) signalling and T cell development. It is expressed in lymphomas, B-cell chronic lymphocytic leukemia, hairy cell leukemia, and acute nonlymphocytic leukemia.

In addition to the SJL, the Ptprca (Ly5.1) allele is also carried by the STS/A and DA strains. The Ptprcb (Ly5.2) allele is expressed by the C3H, DBA/1 et DBA/2, CBA/J and AKR strains as well as the C57BL/6.
Mutation
The recessive a or b mutation (chromosome 1) is a nucleotide substitution: sequencing of Ptprc identified a T to C point mutation resulting in a phenylalanine to serine missense mutation. Twelve nucleotide differences between the a and b alleles have been identified. These base substitutions correspond to five amino-acid changes within the extracellular domain of the encoded protein. These amino-acid differences are clustered in a region that also contains the greatest divergence between mouse and rat sequences.
Homozygotes are viable, fertile, normal in size and do not display any gross physical or behavioral abnormalities.
These mice contain only 15% of the normal level of CD45 expression on the surface of nucleated hematopoietic cells, without altering splicing of transcript isoforms. These mutant mice may be useful in studying the differential regulation of TCR signalling by altered CD45 expression levels.
Homozygous null mutants have defective T cell, B cell, and NK cell morphology and physiology. Mice carrying an engineered point mutation exhibit lymphoproliferation and autoimmunity that leads to premature death.
References
- Fischer et al.1991, Protein tyrosine phosphatases: a diverse family of intracellular and transmembrane enzymes, Science 253: 401-406.
- Charbonneau et al. 1988, The leukocyte common antigen (CD45): a putative receptor-linked protein tyrosine phosphatase, PNAS USA 85(19):7182-6.
- Morse et al. 1987, Genetic nomenclature for loci controlling mouse lymphocyte antigens, Immunogenetics 25(2):71-8.
- Komura et al. 1975, Ly-5: a new lymphocyte antigen system, Immunogenetics 1:452-6.
Cdh23ahl is the mutation responsible for hearing loss in multiple inbred strains and impacts mice hearing phenotypes as a genetic modifier.
Cdh23ahl (cadherin 23 (otocadherin); age related hearing loss 1) is a recessive mutation, on the Cdh23 gene (chromosome 10). Further analysis has shown this is caused by a G to A transition at nucleotide position 753 of Cdh23. This hypomorphic allele causes in frame skipping of exon 7, which is predicted to delete part of the 2nd and 3rd ectodomains, and cause reduced message stability.
A G nucleotide at this site (Cdh23c.753G) results in normal exon splicing, whereas an A nucleotide (Cdh23c.753A) disrupts the canonical donor splice site sequence and causes in-frame exon skipping. The Cdh23c.753G allele is associated with resistance to AHL and is dominant to the recessive Cdh23c.753A allele, which is associated with AHL susceptibility.
It results in progressive hearing loss with onset after 10 months of age in C57BL/6J and C57BL/6N strains and after 3 months of age in BALB/cByJ, DBA/1J and DBA/2J strains.
References
- Johnson et al. 2017, Effects of Cdh23 single nucleotide substitutions on age-related hearing loss in C57BL/6 and 129S1/Sv mice and comparisons with congenic strains, Sci Rep. 2017; 7: 44450
- Kane et al. 2012, Genetic background effects on age-related hearing loss associated with Cdh23 variants in mice, Hear Res. 2012 Jan; 283(1-2): 80–88
- Davis, 2001, Genetic basis for susceptibility to noise-induced hearing loss in mice, Hear Res. 2001 May;155(1-2):82-90
- Di Palma, 2001, Genomic structure, alternative splice forms and normal and mutant alleles of cadherin 23 (Cdh23), Gene. 2001 Dec 27;281(1-2):31-41
- Willott et al. 1995, Genetics of age-related hearing loss in mice. II. Strain differences and effects of caloric restriction on cochlear pathology and evoked response thresholds, Hear Res. 1995 Aug;88(1-2):143-55
Pde6brd1 (phosphodiesterase 6b, retinal degenerescence 1), homozygous for the rd1 mutation have an early onset severe retinal degeneration to the blindness, due to a murine viral insert and a second recessive nonsense mutation in exon 7 of the Pde6b gene (chromosome 5) encoding the beta subunit of cGMP-PDE (enzyme) in the retinal rod photoreceptor. Rod cells are lost by 35 days of age; cone cells degenerate slower and some light sensitivity persists.
The CBA/J, C3H/HeN, SJL/J and FVB/N strains carry the rd1 mutation.



Crb1rd8 (crumbs family member 1, photoreceptor morphogenesis associated; retinal degeneration 8) is a single base deletion in the Crb1 gene on the chromosome 1. This deletion causes a frame shift and a premature stop codon that truncates the transmembrane and cytoplasmic domain of the protein after amino acid 1207. This recessive mutation causes photoreceptor degeneration observed in spots, caused by retinal folds and pseudorosettes with large white retinal deposits covering the inferior quadrant of the retina, found in the fundus of the eye.
The C57BL/6N strain carries the rd8 mutation.
References
- Mattapallil et al. 2012, The Rd8 Mutation of the Crb1 Gene Is Present in Vendor Lines of C57BL/6N Mice and Embryonic Stem Cells, and Confounds Ocular Induced Mutant Phenotypes, Invest Ophthalmol Vis Sci. May; 53(6): 2921–2927
- Aleman et al. 2011, Human CRB1-Associated Retinal Degeneration: Comparison with the rd8 Crb1-Mutant Mouse Model, Invest Ophthalmol Vis Sci. Aug; 52(9): 6898–6910
- Chang et al. 2002, Retinal degeneration mutants in the mouse, Vision Res. 2002 Feb;42(4):517-25
- Giménez and Montoliu, 2001, A simple polymerase chain reaction assay for genotyping the retinal degeneration mutation (Pdeb(rd1)) in FVB/N-derived transgenic mice, Lab Anim. 2001 Apr;35(2):153-6
HOW IS THE PRODUCTION IN UNITS?
Our animals are in a protected environment:
-
-
- The air is filtered in HEPA H13
- The unit is over pressurized (positive pressure)
- All inputs are sterilized: all work suits, material (cages, lids, bottles, racks, etc.) and consumables (bedding, food, enrichment, etc.) are autoclaved
- Incoming flows, internal flows and outflows respect the “go-forward principle”
- The staff enters a barrier through a succession of 5 changing rooms
-

Only the animal staff has access to the units. Staff wears permanently sterile gowning containing: gloves, mask, mob cap (covering completely the hair and the ears) and appropriate work clothing for every single person.
Environmental conditions:
- Temperature and humidity are subject to daily monitoring.
- The light cycle is 12h/12h for rats and mice. This cycle is 14h/10h (day/night) for hamsters.
- The water supplied to animals is acidified to pH 5 and undergoes chlorination (chlorine from 6 to 8 ppm).
- The food distributed in areas of production depends on the strain.
- The bedding (autoclaved) is spruce bedding.
- Enrichment for young (wood bricks to gnaw type Aspen brick) and breeders (strips of unbleached kraft brown paper cut and folded type sizzle nest)


RATS
In our production areas of rats, couplings are in permanent monogamy; the male is left with the female to have litters continuously. To mate, the females are aged at least 10 weeks and males at least 12 weeks of age for outbred rats and 10 weeks of age for the inbred and mutant rats.
Gestation in rats lasts 20 to 23 days. The average litter size varies depending on the strain of rat. Outbred strains of rats (Wistar, Sprague Dawley, Long Evans) generally give more pups per litter.
MICE
In our production areas of mouse, couplings are in permanent bigamy: 2 females with 1 male, left continuously. An exception is made for SJL/J mice with couplings in non-permanent bigamy due to their aggressive behavior. To mate, the females are aged at least 10 weeks and males at least 12 weeks of age for outbred strains and at least 10 weeks of age for inbred mice. SJL/J and NOD-scid mice are mated from at least 7 weeks old. Diabetic mice are coupled from the age of 8 weeks for females and males. For NMRI nude mice, females are mated from 10 weeks of age and males from 8 weeks old.
The duration of gestation in mice is 18-20 days depending on the strain. The average litter size varies with the strain. Outbred strains of mice (NMRI and SWISS) generally give more pups per litter.
HAMSTERS
In our production areas of hamsters, couplings are in non- permanent monogamy due to the aggressive behavior of the female out of the period of estrus. Females are coupled from 8 weeks of age and males from 12 weeks of age.
GERBILS
In our production areas of gerbils, couplings are in permanent monogamy. Females are coupled from the age of 10 weeks and males from 12 weeks of age.
WEANING
For all strains in our facilities, weaning is made at 3 weeks of age (21 days).
At the weaning at 3 weeks old, a sexing allows the separation of males and females. A second sexing at 4 weeks of age ensures non sex mix of animals in cages.
At weaning, males and females are separated. Thus, females from different litters are grouped in a same production cage. It is the same for males.



At Janvier Labs, we have improved our production of obese animals (ob/ob) in order to better meet customer demands while integrating ethics, economics and practice.
Currently, our production of obese ob/ob individuals is by IVF using homozygous ob/ob oocytes and spermatozoa (recipient B6CBA/F1 females) to obtain 100% individuals with ob/ob genotype from birth. This is a major improvement over our prior production with 25% of ob/ob individuals in litters due to heterozygous parent crosses ob/+ due to the infertility problem of homozygous ob/ob individuals.
Several other advantages are brought by this mode of production of obese individuals by IVF:
- We no longer have the need to wait until the age of 5 weeks to know the genotype of individuals by the obese phenotype;
- No need for genotyping to ensure ob/ob genotype (cost gain);
- Only the animals of interest were generated at the request of the client, which avoids the production, the breeding and the euthanasia of animals not requested (ethics with the rule of the 3R and gain of cost);
- There is no longer competition in litters between homozygotes and heterozygotes, leading to better growth and homogeneity of ob/ob individuals shipped to customers;
- Reinforcement of consanguinity by the use of single-father spermatozoa for the entire production of ob/ob individuals (frozen semen from a single ob/ob male placed in contact with oocytes of ob/ob females donor);
- Standardization of the intestinal flora of the animals produced (intestinal flora of recipient B6CBA/F1 animals).
- An ability to also produce controls by IVF with an assured heterozygous genotype ob/+, unlike before where only genotyping could ensure the genotype of controls in litters obtained with ob/+ and +/+ (genotype assurance of obese controls and cost gain).
However, the request of ob/ob animals needs more delay. The customer needs to order animals earlier to have them on the desired date.
For couplings, females are aged from 9 to 12 weeks and males are used from 10 weeks to 5 months (sterility tested). When sorting females for time-mating, females reserved must not show any abnormality. Females are placed with males in monogamy at the end of the day (E -1) (just before, sorting of estrus by impedance measuring for female rats / by vulva observation for female mice) until the next morning (E0) (uncoupling and statement of vaginal plugs).
Vaginal plugs after mating lasts less than 24 hours. For us, the females are put in coupling the night. The next morning at 8 am readings plugs are made to prevent them from falling that occurs in the afternoon. Only females with vaginal plug are kept for sale. However, the presence of this plug (sign of a mating) doesn’t guarantee a 100% pregnancy.
The percentage of successful fertilization with respect to the number of vaginal plugs observed in females after mating is specific to a strain.
For mice, according to our shipping procedures for pregnant females, a visual check is performed on each female to detect pregnancy. Gestation is only guaranteed at E15 for outbred strains and at E16 for inbred strains, the detection of a gestation being more difficult before that date. Thus, it is possible that the lack of pregnancy is not detected when we control some females.
For rats, females are weighed on E0 (day of uncoupling) and on E8 for Wistar female rats and on E11 for SD female rats, to confirm the diagnosis of pregnancy. According to our shipping procedures for pregnant females, a check is performed on each female to detect pregnancy. In the preparation of time-mated females, they are controlled by a visual examination and weighing. A significant weight gain means gestation. However, this weight gain was not related to the numbers of embryos. Gestation is only guaranteed at E15 for outbred strains and at E16 for inbred strains, the detection of a gestation being more difficult before that date. Thus, it is possible that the lack of pregnancy is not detected when we control some females.